Resoconto autunno 2004 - Gruppo Inanellamento Limicoli (GIL, Napoli)
Menu principale:
- Home Page
- L'Associazione
-
Ricerche in atto
- Migrazione autunnale Allodola
- Migrazione primaverile Tordo
- Ali Tordo bottaccio in Campania
- Stazione Monte Vaccaro
- Riserva Naturale Foce Volturno
- Inanellamento Medio Volturno
- Avifauna svernante
- Avifauna Foce Neto e Sila
-
Ricerche concluse
- Ali Turdidi e Beccacce ATC LT 2
- Stazione Malga Rosello
- Migrazione Caradriiformi
- Progetto Rondine
- Migrazione autunnale Tordo
- Ali Turdidi e Beccacce ATC CE
- Progetto Alauda 2000
- Osservatorio Roccolo Ganda
- Atlante Migrazioni Puglia
- Download
- News
Resoconto autunno 2004
MONITORAGGIO DELLA MIGRAZIONE AUTUNNALE NELL'OSSERVATORIO ORNITOLOGICO "DANIELE ANESA",
ROCCOLO GANDA -
Resoconto dell'attività
27 agosto -
Sergio Scebba
Realizzazione dell'iniziativa
U.O. Pianif. Faunistica e Venatoria Regione Lombardia
Progetto
Gruppo Inanellamento Limicoli (G.I.L., Napoli)
Responsabile
Dr. Sergio SCEBBA, Dottore in Medicina e Chirurgia
Studio e Gestione dell'Ambiente Naturale
Viale Augusto 62 -
E-
Collaborazione
Dr. Giuseppe LA GIOIA, Biologo, Dottorato in Ecologia ed Ornitologo
Or.Me. -
Via Cap. Ramirez 89 -
Partecipanti
Cristian ANESA (Italia), Dean BACKHOUSE (Gran Bretagna), Susan BRUNELL (Gran Bretagna), John GLAZEBROOK (Gran Bretagna), Bruce HALLS (Gran Bretagna), Kent HIDAY (Stati Uniti), Amy KAEHN (Stati Uniti), Richard LIVINGTON (Gran Bretagna), Gabor LOVEI (Ungheria), Alan MILLER (Gran Bretagna), Pam NEIMAN (Stati Uniti), Paul NEWTON (Gran Bretagna), Angelo POLI (Italia), Nicole RODNEY (Stati Uniti), Ashish SRIVASTAVA (India), Viviana SABBIONI (Italia), Daniela SCEBBA (Italia), Jelena TELEGINA (Lituania), Olena TRYASKO (Ucraina), Alessio USAI (Italia), Anna VANNUCCHI (Italia), Rodney WEST (Gran Bretagna), Natalino XUEREB (Malta)
*****************************************

Abbiamo cercato di mantenere ancora vivo il tuo Roccolo, anche se limitati da altoparlanti, CD, reti mist-
Ed il Roccolo non è stato più lo stesso.
PREMESSA
Dal 1991 al 2002 l'Osservatorio Ornitologico "Roccolo Ganda" (Aviatico-
Tutti i dati raccolti sono stati riportati su schede prestampate e poi archiviati nella banca-
La possibilità di condurre queste ricerche la si deve al lavoro inestimabile effettuato dal proprietario dell'impianto, l'Architetto Daniele Anesa, che è riuscito a creare una struttura che costituisce un esempio eccezionale di stazione ornitologica per lo studio delle migrazioni. Purtroppo, in seguito alla sua prematura scomparsa, l'Osservatorio non ha funzionato nell'autunno del 2003 e la sua assenza potrebbe determinare la definitiva chiusura di ogni attività; verrebbe quindi meno la possibilità di far conoscere uno dei più importanti ed antichi Roccoli della Lombardia, oltre naturalmente a causare una grave perdita per la ricerca scientifica. Pertanto, il Gruppo Inanellamento Limicoli, Associazione culturale di volontariato senza fini di lucro impegnata in attività di ricerca sull'avifauna selvatica, per non disperdere questo patrimonio e per assicurare una continuità alle ricerche avviate, si propone di gestire l'impianto per quanto riguarda l'attività scientifica e di costituire l'Osservatorio Ornitologico "Daniele Anesa".
In questa relazione vengono riportati i risultati relativi all'attività svolta nell'autunno 2004 quale primo contributo conoscitivo della migrazione autunnale in provincia di Bergamo.

OBIETTIVI DELLO STUDIO
Lo scopo di questo studio è il monitoraggio dei flussi migratori post-
1 -
2 -
3 -
4 -
5 -
6 -
7 -
8 -
9 -
Altri aspetti del progetto
Sulla base di eventuali collaborazioni con altri Istituti scientifici sarà possibile avviare ulteriori ricerche, quali:
-
-
-
-
-
-
AREA DI STUDIO
L'Osservatorio Ornitologico "Daniele Anesa" si trova in località Ganda (45°47'N 09°48'E) nel comune di Aviatico, provincia di Bergamo, ad un'altezza di 1034 metri sul livello del mare. L'impianto, situato nel distretto geobotanico prealpino delle Alpi Orobie, è collocato a ridosso della linea di spartiacque che separa la Valle Seriana dalla Valle Brembana, ed è posizionato in prossimità del limite superiore del bosco, in modo tale da creare un continuum con la vegetazione circostante. Infatti il Roccolo ha una sua composizione vegetazionale tipica che ben si associa con l'habitat naturale. L'area è caratterizzata da un suolo di tipo argilloso, con piccoli affioramenti di roccia calcarea, che favorisce la crescita e lo sviluppo delle piante tipiche di questa fascia vegetazionale.

La Valle Seriana vista dall'Osservatorio Ornitologico
Il bosco si presenta come una associazione tra Querce e Faggio, dove il Faggio Fagus sylvatica è l'essenza dominante. Associate a questa pianta sono il Carpino bianco Carpinus betulus e il Frassino Fraxinus excelsior. A quote più basse, la composizione vegetazionale arborea subisce una mescolanza con altre specie quali Castagno Castanea sativa, Farnia Quercus robur e Rovere Quercus petraea. Tale distribuzione altitudinale rispecchia, in linee generali, i limiti superiore ed inferiore dei rispettivi piani, basale e montano, delle fasce vegetazionali alpine e prealpine. Ai margini del bosco, ma anche nelle parti interne, sono presenti ed abbondanti essenze da frutto e da bacca. Il Sambuco Sambucus nigra e l'Agrifoglio Ilex aquifolium sono tra le piante più frequenti e dominanti nel sottobosco della faggeta; esse sono presenti nelle forme arbustive ed arboree insieme a cespugli di Rovo Rubus fruticosus, a piante di Sorbo montano Sorbus aria ed a formazioni rampicanti di Edera Hedera helix. La presenza delle attività antropiche ha dato forma e composizione alla vegetazione boschiva: sulle vette circostanti sono presenti dei rimboschimenti ad Abete (Abete rosso Picea excelsa) e non manca una certa diversità arborea nei dintorni delle abitazioni che concorre ad incrementarne la diversità (soprattutto piante quali Noce Juglans regia, Nocciolo Corylus avellana e Alloro Laurus nobilis); inoltre, laddove viene ancora praticato il pascolo di ovini e bovini, si ha la formazione di praterie. Il lento abbandono della pratica dell'alpeggio in questo territorio sta portando alla scomparsa dei pascoli alpini a bassa quota a vantaggio di aggruppamenti forestali mesofili di caducifoglie.

La composizione vegetazionale del Roccolo si discosta poco da quella del bosco. L'essenza dominante è il Carpino bianco che, insieme col Faggio, costituisce l'impalcatura su cui si regge l'intero impianto; queste piante ben si prestano al taglio e al modellamento secondo le esigenze culturali e tradizionali di gestione di tali strutture, formando i corridoi in cui correranno le reti per la cattura degli uccelli. Le altre piante presenti, che si sviluppano in forma arborea, sono tipiche essenze da bacca. La più caratteristica è il Sorbo degli Uccellatori Sorbus aucuparia con frutti che nella stagione autunnale si presentano di colore rosso, portati in dense infruttescenze a corimbo.
Le altre essenze sono: l'Agrifoglio con foglie sempreverdi e frutti di colore rosso vivace alla maturità, il Biancospino Crataegus oxyacanta con frutti che sono piccole drupe di colore rosso, il Ginepro comune Juniperus communis con piccoli frutti di colore nero-

Sorbo degli Uccellatori Sorbus aucuparia
La composizione vegetazionale qui descritta riflette le condizioni climatiche che si presentano in quest'area. Inoltre, data la conformazione geografica della valle e la presenza del fiume Serio che scorre sul fondo della stessa, si vengono a creare ulteriori microclimi che influenzano la disposizione delle varie comunità vegetali. Il bosco che si estende avanti al Roccolo è posizionato tra due versanti montuosi che ne limitano il periodo di esposizione alla luce e lo proteggono dai forti venti. Si instaura, in tal modo, un microclima che favorisce la crescita di piante tipicamente mesofile cioè che non sopportano in misura eccessiva né il freddo né il caldo, né l'umidità né la secchezza, né l'ombra né la luce, e il Faggio ne è l'esponente principale in quest'area.
Le condizioni climatiche della zona sono riconducibili a una forma di transizione tra il clima continentale, tipico della pianura padana e quello di tipo alpino; il clima anemologico è influenzato dalle stagioni e con direzione preferenziale di scorrimento del vento lungo l'asse NE-
METODOLOGIE
La cattura e l'inanellamento degli uccelli, tra le varie metodiche utilizzate per studi sull'avifauna, rappresenta quella di più comune impiego, in quanto offre un utile mezzo per monitorare i flussi migratori e verificare l'entità della presenza delle diverse specie nel corso della migrazione. La cattura permette di effettuare un esame ravvicinato di ciascun individuo e quindi di rilevare una serie di dati, quali età, sesso, biometria, muta, peso, accumuli di grasso, nonché qualsiasi altra caratteristica si ritenga necessaria. Inoltre questa metodica, mediante l'apposizione di un anello numerato in maniera univoca, consente di conoscere in caso di ritrovamento, la località esatta in cui l'uccello è stato preso per la prima volta ed il tempo trascorso dal momento del suo inanellamento. Queste informazioni consentono di determinare le rotte seguite dai migratori e di analizzarne la distribuzione geografica e temporale, di individuare le aree di nidificazione e di svernamento delle diverse popolazioni, di studiare le correlazioni che esistono tra la migrazione e le condizioni del tempo (passaggi di aria fredda, influenza della velocità del vento e della direzione, pressione barometrica, inverni con ondate di freddo particolarmente intenso) e di valutare il diverso comportamento dei giovani e degli adulti.

Inanellamento di un Tordo bottaccio con anelli in alloy dell'Istituto Nazionale Fauna Selvatica
Il monitoraggio della migrazione autunnale è stato quindi basato su una costante e standardizzata attività di cattura e inanellamento: per tutto il periodo della migrazione la metodologia di cattura è stata standardizzata mediante l'utilizzo di reti verticali tipo mist-
Dopo la cattura, gli uccelli sono stati posti in sacchetti di tela per evitare che potessero ferirsi e quindi subito inanellati con anelli dell'Istituto Nazionale Fauna Selvatica applicati al tarso. Su tutti gli uccelli catturati è stata identificata l'età ed il sesso (Svensson 1992, Baker 1993); per codificare le classi d'età sono stati utilizzati i rispettivi codici EURING:
età 2, esatto anno di nascita sconosciuto;
età 3, nato nell'anno in corso;
età 4, nato prima dell'anno in corso, ma l'esatto anno di nascita è sconosciuto.
Sono state rilevate di routine alcune misure biometriche: lunghezza dell'ala (misurata secondo il metodo della corda massima, appiattita e raddrizzata, Svensson 1992) con approssimazione di 0,5 mm; lunghezza della III remigante primaria (massima lunghezza, Berthold & Friedrich 1979) con approssimazione di 0,5 mm; lunghezza del tarso (misurata con un calibro, Svensson 1992) con approssimazione di 0,1 mm. È stato inoltre rilevato il peso mediante l'uso di una bilancina elettronica con approssimazione di 0,1 gr. ed è stata effettuata una stima della quantità di grasso visibile, attribuendo un punteggio variabile da 0 a 8 in base alla quantità presente alla furcula e sull'addome (Kaiser 1993). E' stato controllato lo stato della muta delle penne del corpo usando una scala da 0 a 2 (0, nessuna penna in crescita; 1, fino a 20 penne in crescita; 2, più di 20 penne in crescita). Inoltre, per alcune specie è stata misurata l'apertura e la superficie alare. La prima corrisponde alla massima distanza che intercorre tra le punte delle due ali con le articolazioni completamente distese con approssimazione di 1 mm, la seconda all'area delle due ali più la parte del corpo compresa tra l'attaccatura delle ali proiettate su di una superficie piana (Pennycuick 1999). La raccolta di questi dati consentirà di calcolare, mediante la versione 1.11 (Marzo 2002) per Windows del programma "Flight" di Pennycuick, l'autonomia di volo e la sua durata (definita come il tempo necessario per percorrere le distanze predette) in assenza di vento di ciascun uccello.

Misurazione della III remigante primaria
Le operazioni di inanellamento, identificazione dell'età e del sesso, raccolta delle misure biometriche sono state effettuate da inanellatori esperti autorizzati dall'INFS, coadiuvati da alcuni aspiranti inanellatori e collaboratori del Gruppo Inanellamento Limicoli. L'intera procedura è durata circa 60-
L'attività di cattura ed inanellamento è stata effettuata continuativamente dal 27 agosto al 27 novembre 2004 (pentadi 49-
Tutti i dati inerenti l'inanellamento sono stati quindi riportati su schede prestampate e successivamente archiviati in un Data-
L'andamento della migrazione è stato descritto mediante periodi standard di cinque giorni secondo lo schema per pentadi (Berthold 1973) con la prima pentade che va dal 1 al 5 gennaio e l'ultima, la 73, che va dal 27 al 31 dicembre. Anche se brevi, questi spazi temporali presentano un potere risolutivo sufficiente per evidenziare le diverse fasi dell'andamento migratorio (inizio, picco, conclusione); inoltre l'unità di misura adottata, in quanto sottomultiplo del numero di giorni di un anno solare, è esattamente replicabile negli anni successivi. Ciò consente di confrontare campioni di dati raccolti in periodi analoghi ma provenienti da anni o aree differenti.
L'analisi dell'indice di flusso (numero di uccelli inanellati per superficie di reti moltiplicato per ore di cattura) per un periodo sufficientemente lungo, potrà inoltre consentire lo studio della dinamica di popolazione, oltre che di stimare l'entità dei flussi nell'area.
RISULTATI E DISCUSSIONE
Analisi delle catture
La stagione di studio è stata realizzata con 93 giornate di catture effettive, che hanno prodotto una quota media di 44 uccelli catturati per ciascuna giornata con un valore minimo di due catture il 27 ottobre e l'1 novembre ed un valore massimo di 154 il 5 ottobre. In totale sono stati catturati 4.108 uccelli appartenenti a 56 specie diverse; di queste, 4 sono non-
Va rimarcata la notevole differenza con i risultati ottenuti negli anni precedenti di attività del Roccolo Ganda: nel 2001 sono state effettuate 12.802 catture (di cui 3.683 Pettirossi) e nel 1997 si è addirittura arrivati a 13.316 (di cui 3.604 Lucarini). L'elenco completo delle catture del 2004 è presentato in tabella 1.
Tabella 1. Elenco delle specie inanellate nel periodo 27 agosto/27 novembre 2004. |
|||||
Specie |
Agosto |
Settembre |
Ottobre |
Novembre |
TOT |
Sparviere - |
- |
4 |
1 |
- |
5 |
Assiolo - |
- |
1 |
- |
- |
1 |
Torcicollo - |
1 |
6 |
1 |
- |
8 |
Picchio rosso maggiore - |
- |
- |
- |
1 |
1 |
Rondine - |
1 |
19 |
- |
- |
20 |
Prispolone - |
23 |
144 |
4 |
- |
171 |
Pispola - |
- |
- |
1 |
- |
1 |
Spioncello - |
- |
- |
- |
2 |
2 |
Ballerina gialla - |
- |
2 |
- |
- |
2 |
Ballerina bianca - |
2 |
1 |
- |
- |
3 |
Scricciolo - |
- |
1 |
6 |
- |
7 |
Passera scopatola - |
- |
- |
15 |
1 |
16 |
Pettirosso - |
10 |
762 |
936 |
26 |
1734 |
Codirosso spazzacamino - |
- |
2 |
13 |
2 |
17 |
Codirosso - |
14 |
98 |
17 |
- |
129 |
Stiaccino - |
2 |
2 |
- |
- |
4 |
Saltimpalo - |
- |
2 |
1 |
- |
3 |
Merlo - |
2 |
3 |
14 |
3 |
22 |
Cesena - |
- |
- |
- |
43 |
43 |
Tordo bottaccio - |
- |
47 |
78 |
3 |
128 |
Tordo sassello - |
- |
- |
5 |
3 |
8 |
Tordela - |
- |
- |
3 |
3 |
6 |
Forapaglie macchiettato - |
- |
1 |
- |
- |
1 |
Cannaiola - |
- |
3 |
- |
- |
3 |
Canapino maggiore - |
- |
2 |
- |
- |
2 |
Bigiarella - |
- |
3 |
- |
- |
3 |
Sterpazzola - |
- |
1 |
- |
- |
1 |
Beccafico - |
4 |
142 |
2 |
- |
148 |
Capinera - |
8 |
122 |
45 |
- |
175 |
Luì bianco - |
1 |
- |
- |
- |
1 |
Luì verde - |
- |
3 |
- |
- |
3 |
Luì piccolo - |
- |
7 |
29 |
- |
36 |
Luì grosso - |
4 |
20 |
- |
- |
24 |
Regolo - |
- |
3 |
27 |
8 |
38 |
Fiorrancino - |
- |
2 |
1 |
- |
3 |
Pigliamosche - |
- |
3 |
- |
- |
3 |
Balia nera - |
130 |
265 |
1 |
- |
396 |
Codibugnolo - |
2 |
- |
1 |
5 |
8 |
Cincia mora - |
1 |
10 |
1 |
6 |
18 |
Cinciarella - |
6 |
17 |
28 |
7 |
58 |
Cinciallegra - |
6 |
- |
10 |
15 |
31 |
Picchio muratore - |
- |
- |
2 |
- |
2 |
Rampichino alpestre - |
- |
- |
- |
2 |
2 |
Averla piccola - |
1 |
4 |
- |
- |
5 |
Ghiandaia - |
2 |
- |
- |
- |
2 |
Passera d'Italia - |
- |
3 |
- |
- |
3 |
Fringuello - |
2 |
17 |
392 |
188 |
599 |
Peppola - |
- |
- |
42 |
94 |
136 |
Verdone - |
2 |
- |
- |
- |
2 |
Cardellino - |
26 |
8 |
1 |
9 |
44 |
Lucarino - |
- |
1 |
15 |
1 |
17 |
Frosone - |
- |
- |
5 |
2 |
7 |
Zigolo giallo - |
1 |
2 |
- |
- |
3 |
Zigolo muciatto - |
- |
- |
- |
1 |
1 |
Ortolano - |
- |
1 |
- |
- |
1 |
Migliarino di palude - |
- |
- |
1 |
- |
1 |
TOTALI |
251 |
1734 |
1698 |
425 |
4108 |
L'andamento generale dell'attività è rappresentato in figura 1, basato sulla percentuale degli uccelli catturati in ciascuna pentade rispetto al numero complessivo delle catture. È possibile evidenziare la presenza di due picchi: il primo, in corrispondenza della pentade 50 (3-
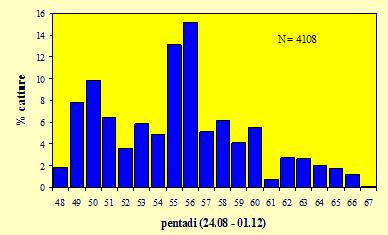
Figura 1. Andamento complessivo delle catture per pentadi nel periodo 27 agosto/27 novembre 2004.
L'attività di cattura standardizzata ha consentito di evidenziare rispetto agli anni precedenti un drastico calo sia del numero complessivo delle catture che di quello di alcune specie, soprattutto Fringillidi, in particolare Lucarino, Peppola e Frosone, e Tordo bottaccio e Tordo sassello. La diversa metodologia utilizzata nell'attuale stagione, reti mist-
La rappresentazione grafica delle percentuali delle specie maggiormente inanellate è mostrata nella figura 2.
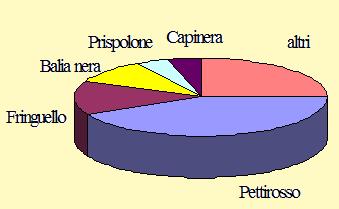
Figura 2. Percentuale delle specie inanellate nel periodo 27 agosto/27 novembre 2004 (n = 4.108).
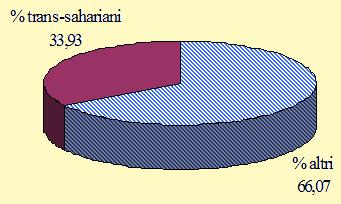
Figura 3. Migratori su lunga e media distanza: percentuale totale delle specie inanellate nel periodo 27 agosto/27 novembre 2004 (n = 56).
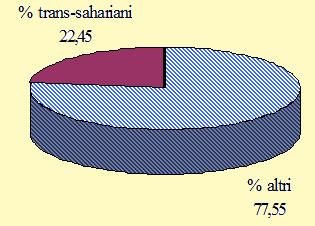
Figura 4. Migratori su lunga e media distanza: percentuale totale degli uccelli catturati nel periodo 27 agosto/27 novembre 2004 (n = 4.108).
L'attività di ricerca, condotta per un ampio arco di tempo, dalla fine di agosto alla fine di novembre, ha consentito di descrivere, in termini generali, le principali variazioni nella composizione dei flussi degli uccelli che attraversano le Alpi in differenti momenti della migrazione post-
Analisi della migrazione delle specie più rappresentative
Per effetto delle profonde trasformazioni ambientali, dell'inquinamento e di tutte le altre forme di impatto antropico, si è verificata una drammatica riduzione delle aree utilizzate per la sosta durante le migrazioni e poi per lo svernamento. Per predisporre adeguati piani di conservazione risulta di fondamentale importanza raccogliere tutte le informazioni attualmente disponibili; d'altra parte i cambiamenti politici avvenuti nell'Europa dell'Est ed il recente ingresso nella Comunità Europea di alcuni di questi paesi consentono migliori opportunità per coordinare a livello internazionale opportune strategie di conservazione e per sviluppare uno scambio di dati e di cooperazione.
La Valle Seriana e l'area del Roccolo Ganda in particolare, come è stato dimostrato da questa ricerca e dai dati raccolti negli anni precedenti, rappresentano un'importante zona di passaggio durante la migrazione post-
L'attività svolta nel corso di questo anno ha fatto registrare una drastica contrazione del numero delle catture rispetto a quelli invece ottenuti negli anni precedenti. Sicuramente la migrazione autunnale è stata fortemente condizionata dalle condizioni climatiche; il succedersi di continue perturbazioni dalla seconda decade di ottobre alla prima di novembre con relative abbondanti precipitazioni, potrebbe aver determinato localmente condizioni poco favorevoli per il passaggio e la sosta, nonché particolari condizioni meteorologiche sull'arco alpino in grado di influire sul transito dei migratori in movimento verso le aree di svernamento. La presenza di aree di bassa pressione potrebbe aver spinto gli uccelli sia ad utilizzare rotte e punti di sosta differenti sia a sorvolare la zona senza fermarsi. Ciò potrebbe aver determinato l'evidente fluttuazione del numero degli individui osservati che si è poi riflessa anche sul numero delle catture effettuate. Infatti, rispetto all'attività svolta nel 2002 si è verificato un calo del 61% che arriva addirittura quasi al 68% se il confronto viene fatto con quella del 2001. Queste flessioni possono comunque rientrare nel normale andamento della fenologia della migrazione in quanto, da un anno all'altro, è possibile che si verifichino delle grosse variazioni nei passaggi dei migratori. D'altra parte uno degli scopi di questa ricerca è proprio quello di monitorare l'andamento della migrazione nel corso degli anni. L'area è stata comunque utilizzata da un elevato numero di specie, tra cui le più catturate sono state il Prispolone tra i Motacillidi, il Pettirosso, il Codirosso ed il Tordo bottaccio tra i Turdidi, il Beccafico e la Capinera tra i Silvidi, la Balia nera tra i Muscicapidi, il Fringuello e la Peppola tra i Fringillidi. Per queste nove specie, selezionate in base al numero degli individui catturati, sono stati riassunti i dati sulla fenologia della migrazione e sulla biometria. I grafici mostrano il numero di individui catturati in ciascuna pentade e, indicata da una freccia, la pentade mediana di passaggio calcolata durante il periodo di studio. I dati biometrici riportati nelle tabelle sono relativi alla lunghezza dell'ala, della terza remigante primaria ed al peso ed indicano la media, la deviazione standard e l'entità del campione per ciascuna pentade; inoltre, per Pettirosso e Fringuello, considerato l'elevato numero di dati raccolti, sono riportate anche le distribuzioni di frequenza delle misure della lunghezza dell'ala e del tarso. Un breve testo riassume informazioni generali e commenti sull'andamento della migrazione e sulla biometria; sia nei testi che nelle tabelle le lunghezze dell'ala e della terza sono indicate in millimetri, il peso è dato in grammi.
Prispolone Anthus trivialis
Geonemia
Specie a corologia euroasiatica, il Prispolone nidifica diffusamente in Europa, ad eccezione di Islanda, Spagna centro-

Fenologia della migrazione
Le catture sono state effettuate tra il 28 agosto ed il 17 ottobre 2004 (pentadi 48-
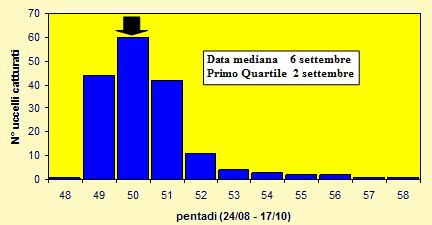
Figura 5. Andamento delle catture per pentade, nel periodo 28 agosto -
Analisi biometrica
In tabella 2 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
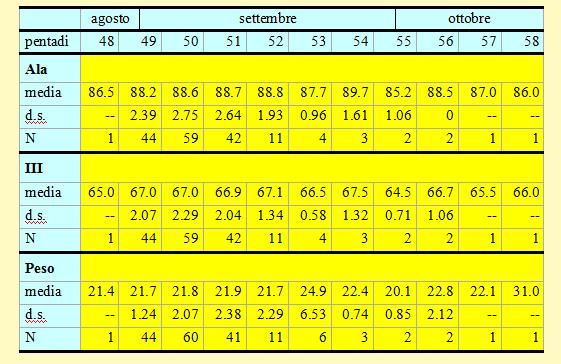
Tabella 2. Misure biometriche (in mm) e peso (in g) dei Prispoloni catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 85.2 a 89.7mm (min 83.0 max 94.5), mentre la media calcolata per l'intero campione è di 88.46mm (d.s. 2.51, n = 170), simile a quella riportata in Cramp (1988) per la Svezia (88.4mm, d.s. 2.69, n = 109). Le medie della III remigante primaria calcolate per ciascuna pentade variano da 64.5 a 67.5mm (min 62.0 max 72.0), mentre la media calcolata per l'intero campione è di 66.91mm (d.s. 2.04, n = 170). Per entrambi le variabili nel corso del periodo di studio non è possibile evidenziare un particolare andamento ed inoltre tra gli uccelli catturati prima e dopo la data mediana delle catture, che occorre nella pentade 50, non si osservano differenze significanti.
I pesi medi calcolati per ciascuna pentade variano da 20.1 a 24.9g (min 17.8 max 31.0), mentre il peso medio totale calcolato per l'intero campione è di 21.85g (d.s. 2.06, n = 170). Nelle pentadi 49-
Analisi del grasso
I pesi medi corrispondenti a ciascuna classe di grasso sono riportati nella tabella 3. I valori rilevati nel corso di questa stagione confermano l'esistenza di una stretta correlazione tra il peso degli uccelli e la quantità di grasso accumulata stimata visualmente.
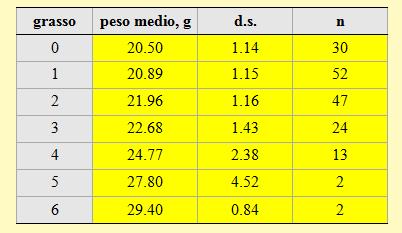
Tabella 3. Peso medio per ciascuna classe di grasso dei Prispoloni catturati nell'Osservatorio ornitologico "Daniele Anesa", autunno 2004.
Va evidenziato che la percentuale di individui che non presentano depositi di grasso visibili o presenti solo in quantità molto ridotta, e quindi inclusi nelle classi di grasso più basse (classe 0 ed 1) costituisce il 48% del campione totale; questa percentuale arriva al 76% se vengono considerati anche gli individui inclusi nella classe 2, cioè con modesti depositi di grasso. Questa situazione conferma quanto è già stato rilevato in precedenza per le medie dei pesi per pentade; pertanto, si può ipotizzare che le riserve di energia non sono sufficienti per raggiungere direttamente i quartieri di svernamento localizzati nell'Africa occidentale e che gli uccelli preferiscono adottare una strategia migratoria caratterizzata da soste intermedie prima di affrontare barriere particolarmente impegnative.
Pettirosso Erithacus rubecula
Geonemia
Specie a corologia europea, il Pettirosso è ampiamente diffuso in Europa dal Mediterraneo alle latitudini più elevate; migratore a corto raggio, sverna soltanto nelle regioni più occidentali e meridionali. In Lombardia è parzialmente sedentario, migratore regolare e svernante a seconda delle popolazioni e nidifica in maniera pressoché uniforme negli ambienti adatti montani e collinari fino a 1700-

Fenologia della migrazione
Le catture sono state effettuate tra il 27 agosto ed il 23 novembre 2004 (pentadi 48-
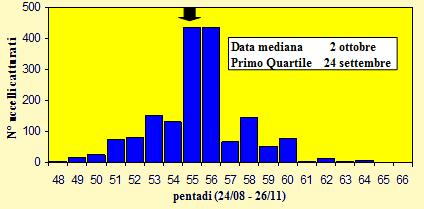
Figura 6. Andamento delle catture per pentade, nel periodo 27 agosto -
Le catture sono distribuite dall'inizio di settembre (pentade 49) alla fine di ottobre (pentade 60) ma presentano una forte concentrazione tra il 28 settembre ed il 7 ottobre (pentadi 55 e 56), periodo in cui è stato realizzato il 50% delle catture totali (869); dalla fine di ottobre alla fine di novembre si registra il passaggio solo di pochi individui: tra le pentadi 61-
Analisi biometrica
In tabella 4 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
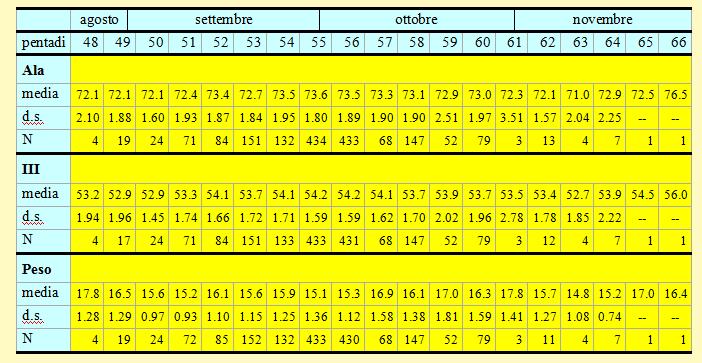
Tabella 4. Misure biometriche (in mm) e peso (in g) dei Pettirossi catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 71.0 a 73.6mm (min 67.5 max 79.0), mentre la media calcolata per l'intero campione è di 73.27mm (d.s. 1.93, n = 1728). Le medie della III remigante primaria calcolate per ciascuna pentade variano da 52.7 a 54.2mm (min 49.0 max 59.0), mentre la media calcolata per l'intero campione è di 53.99mm (d.s. 1.69, n = 1723). Per entrambi le variabili è possibile osservare un incremento dei valori a partire dalla pentade 51; questo potrebbe essere dovuto o ad una migrazione differenziale tra i sessi (le femmine con ala più piccola tenderebbero a passare per prima) oppure, più probabilmente, al passaggio di popolazioni settentrionali di dimensioni maggiori.
I pesi medi calcolati per ciascuna pentade variano da 14.8 a 17.8g (min 12.8 max 21.8), mentre il peso medio totale calcolato per l'intero campione è di 15.64g (d.s. 1.34, n = 1718). Questo valore risulta decisamente inferiore rispetto a quelli calcolati in autunno presso Pisa (Ioalé & Benvenuti 1982) e sull'isola di Vivara presso Napoli (Lövei et al. 1984). Nelle pentadi 53-
Le figure 7 e 8 mostrano le distribuzioni di frequenza, rispettivamente, delle misure della lunghezza dell'ala e del tarso.
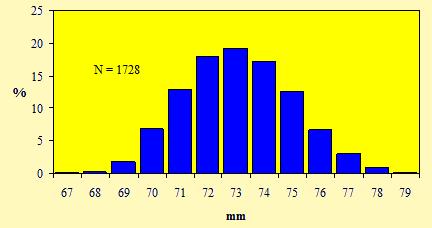
Figura 7. Distribuzione di frequenza delle misure della lunghezza dell'ala dei Pettirossi catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.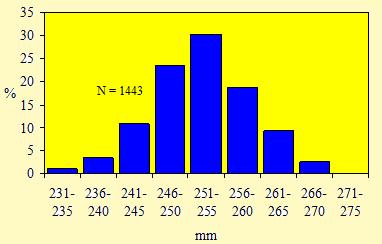
Figura 8. Distribuzione di frequenza delle misure della lunghezza del tarso dei Pettirossi catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Una chiara unimodalità emerge dalle due figure, per cui le lunghezze dell'ala e del tarso non possono costituire un carattere discriminante per determinare il sesso in questa specie. Inoltre questo andamento normale, cioè caratterizzato da una curva a campana con un unico picco, non permette di riconoscere nel campione, oltre che il sesso, anche l'eventuale presenza di individui appartenenti a due o più popolazioni distinte, originarie di regioni differenti.
Occorre tenere presente che, dall'esame delle ricatture di uccelli inanellati in Europa, la Lombardia è interessata da popolazioni provenienti sia dall'Europa settentrionale (Scandinavia, Finlandia, area baltica) che centro-
Codirosso Phoenicurus phoenicurus
Geonemia
Specie a corologia euroasiatica, il Codirosso è ampiamente diffuso in tutta l'Europa fino alle latitudini più elevate esclusa l'Irlanda, l'Islanda e le aree con tipica vegetazione mediterranea; migratore a lungo raggio, sverna principalmente nella savana sub-

Fenologia della migrazione
Le catture sono state effettuate tra il 27 agosto ed il 22 ottobre 2004 (pentadi 48-
A differenza di altre specie di migratori su lunga distanza, quali Prispolone, Beccafico e Balia nera, il Codirosso non presenta le catture concentrate in un determinato periodo; queste sono distribuite abbastanza uniformemente durante quasi tutto l'arco della migrazione, anche se dalla figura 9 è comunque possibile individuare due picchi di maggiore afflusso.
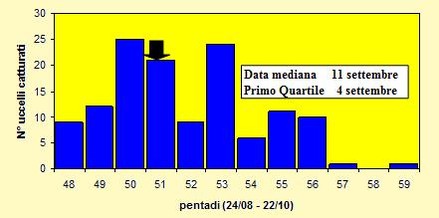
Figura 9. Andamento delle catture per pentade, nel periodo 27 agosto -
Analisi biometrica
In tabella 5 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
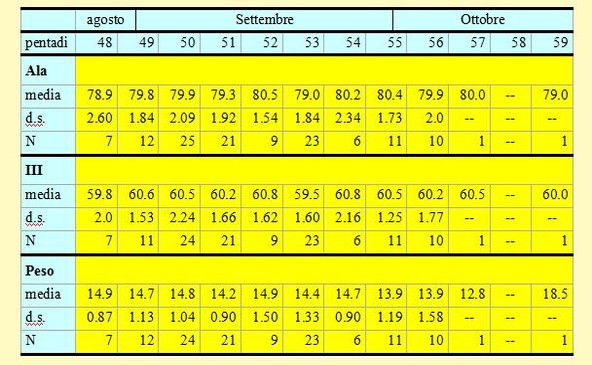
Tabella 5. Misure biometriche (in mm) e peso (in g) dei Codirossi catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 78.9 a 80.5mm (min 74.0 max 84.5), mentre la media calcolata per l'intero campione è di 79.68mm (d.s. 1.95, n = 126). Le medie della III remigante primaria calcolate per ciascuna pentade variano da 59.5 a 60.8mm (min 56.5 max 65.0), mentre la media calcolata per l'intero campione è di 60.26mm (d.s. 1.78, n = 124). Per entrambi le variabili nel corso del periodo di studio non si evidenzia nessuna marcata tendenza; è solo possibile notare che dopo la prima pentade ed in particolare per quanto riguarda la terza primaria, i valori ottenuti non presentano differenze significanti.
I pesi medi calcolati per ciascuna pentade variano da 13.9 a 14.9g (min 12.3 max 18.5), mentre il peso medio totale calcolato per l'intero campione è di 14.49g (d.s. 1.24, n = 125). Nelle pentadi 48-
Analisi del grasso
I pesi medi corrispondenti a ciascuna classe di grasso sono riportati nella tabella 6. I valori rilevati confermano l'esistenza di una buona correlazione tra il peso degli uccelli e la quantità di grasso accumulata stimata visualmente.
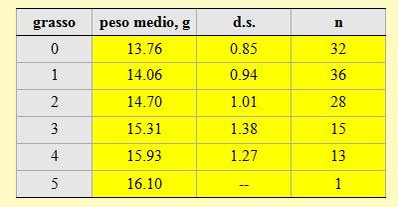
Tabella 6. Peso medio per ciascuna classe di grasso dei Codirossi catturati nell'Osservatorio ornitologico "Daniele Anesa", autunno 2004.
Va evidenziato che la percentuale di individui che non presentano depositi di grasso visibili o presenti solo in quantità molto ridotta, e quindi inclusi nelle classi di grasso più basse (classe 0 ed 1) costituisce il 54% del campione totale; questa percentuale arriva al 77% se vengono considerati anche gli individui inclusi nella classe 2, cioè con modesti depositi di grasso. Questa situazione, analogamente a quanto già riportato per il Prispolone, fa ipotizzare che le riserve energetiche non sono sufficienti per raggiungere direttamente le aree di svernamento localizzate nell'Africa sub-
Tordo bottaccio Turdus philomelos
Geonemia
Specie a corologia eurosibirica, il Tordo bottaccio è uniformemente distribuito come nidificante in tutte le aree boscate dell'Europa centrale e settentrionale, Siberia occidentale ed Asia, mentre nel sud del continente è presente soprattutto sui rilievi; migratore a medio e corto raggio con popolazioni parzialmente o completamente migratrici, sverna nella parte centro-

Fenologia della migrazione
Le catture sono state effettuate tra l'8 settembre ed il 15 novembre 2004 (pentadi 51-
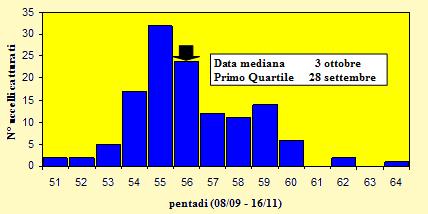
Figura 10. Andamento delle catture per pentade, nel periodo 8 settembre -


Misurazione della lunghezza dell'ala e del tarso
Analisi biometrica
In tabella 7 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
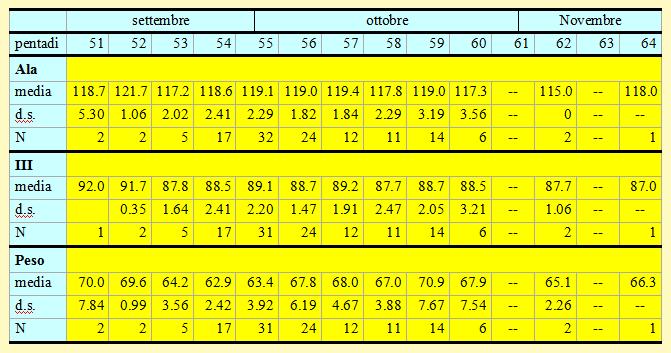
Tabella 7. Misure biometriche (in mm) e peso (in g) dei Tordi bottacci catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 115.0 a 121.7mm (min 112.5 max 125.0), mentre la media calcolata per tutti gli uccelli catturati nel periodo di studio è di 118.73mm (d.s. 2.44, n = 128). Risulta evidente che nelle pentadi 54, 55, 56 e 57, in cui i campioni raccolti sono di una certa entità per il passaggio di un forte flusso migratorio, le differenze tra le medie presentano variazioni minime e quindi non significative. Analoga considerazione può essere fatta anche per le medie della III remigante primaria, le quali variano da 87.7 a 91.7mm (min 82.0 max 95.0), mentre la media calcolata per l'intero campione è di 88.75mm (d.s. 2.13, n = 126).
La lunghezza alare media risulta simile a quella rilevata nel periodo 1997-
I pesi medi calcolati per ciascuna pentade variano da 62.9 a 70.9g (min 56.1 max 83.4); nelle prime fasi della migrazione, cioè dal 18 settembre al 2 ottobre (pentadi 53-

Il sottotondo
Beccafico Sylvia borin
Geonemia
Specie a corologia eurosibirica, il Beccafico in Europa è distribuito principalmente nelle regioni boreali, temperate ed atlantiche, mentre manca completamente nelle aree a clima tipicamente mediterraneo; migratore su lunga distanza, sverna nell'Africa tropicale e meridionale raggiungendo il Sudafrica. In Lombardia è specie estiva e migratrice regolare, nidificante in tutto il settore alpino e prealpino e nelle aree più elevate dell'Appennino pavese, in una fascia altitudinale molto ampia compresa tra i 60-

Fenologia della migrazione
Le catture sono state effettuate tra il 28 agosto ed il 2 ottobre 2004 (pentadi 48-
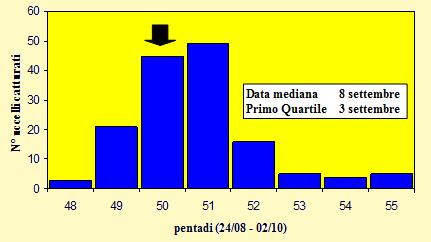
Figura 11. Andamento delle catture per pentade, nel periodo 28 agosto -
Analisi biometrica
In tabella 8 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
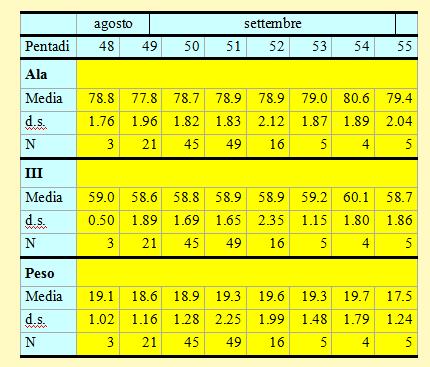
Tabella 8. Misure biometriche (in mm) e peso (in g) dei Beccafichi catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 77.8 a 80.6mm (min 74.0 max 83.5), mentre la media calcolata per l'intero campione è di 78.74mm (d.s. 1.91, n = 148). Le medie della III remigante primaria calcolate per ciascuna pentade variano da 58.6 a 60.1mm (min 54.0 max 63.0), mentre la media calcolata per l'intero campione è di 58.85mm (d.s. 1.75, n = 148). A partire dai primi di settembre (pentade 50), con l'inizio del periodo principale della migrazione, si può osservare un andamento crescente nel corso della stagione dei valori medi della lunghezza alare. Questo potrebbe essere spiegato con la presenza nelle fasi iniziali di individui locali, a cui fa poi seguito il passaggio di popolazioni di dimensioni maggiori probabilmente più settentrionali.
I pesi medi calcolati per ciascuna pentade variano da 17.5 a 19.7g (min 15.4 max 30.2), mentre il peso medio totale calcolato per l'intero campione è di 19.08g (d.s. 1.76, n = 148). Questi valori risultano decisamente inferiori rispetto alla media calcolata in autunno sull'isola di Vivara presso Napoli (22.7g, d.s. 3.7, n = 216, Scebba et al. 1991) e sicuramente sono insufficienti per intraprendere voli su lunga distanza.
Analisi del grasso
I pesi medi corrispondenti a ciascuna classe di grasso sono riportati nella tabella 9. Anche per questa specie i valori rilevati nel corso della stagione confermano l'esistenza di una stretta correlazione tra il peso degli uccelli e la quantità di grasso accumulata stimata visualmente.
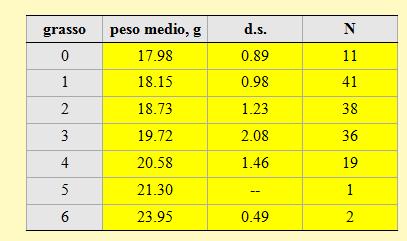
Tabella 9. Peso medio per ciascuna classe di grasso dei Beccafichi catturati nell'Osservatorio ornitologico "Daniele Anesa", autunno 2004.
Rispetto alle altre due specie di migratori trans-
Capinera Sylvia atricapilla
Geonemia
Specie a corologia olopaleartica, la Capinera è ampiamente diffusa in tutta l'Europa, dal Sud della Scandinavia fino alle isole del Mediterraneo, comprese le più piccole. Il comportamento migratorio presenta un'ampia variabilità su base geografica con popolazioni nordiche migratrici su lunga distanza, mentre quelle meridionali sono parzialmente migratrici e/o residenti. Le principali aree di svernamento delle popolazioni europee sono distribuite intorno al bacino del Mediterraneo, dall'Africa settentrionale fino a sud nelle regioni sub-

Fenologia della migrazione
Le catture sono state effettuate tra il 27 agosto ed il 25 ottobre 2004 (pentadi 48-
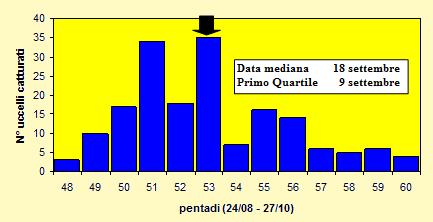
Figura 12. Andamento delle catture per pentade, nel periodo 27 agosto -
Dalla figura 12 si rileva che le catture non sono concentrate in un determinato periodo ma sono abbastanza distribuite durante quasi tutto l'arco della migrazione, anche se nelle pentadi 51 e 53 si possono individuare due picchi di maggiore afflusso.
Analisi biometria
In tabella 10 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
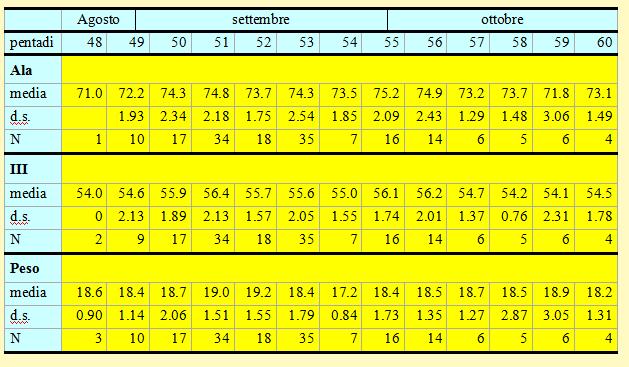
Tabella 10. Misure biometriche (in mm) e peso (in g) delle Capinere catturate nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 71.8 a 75.2mm (min 69.0 max 80.0), mentre la media calcolata per l'intero campione è di 74.14mm (d.s. 2.32, n = 173). Le medie della III remigante primaria calcolate per ciascuna pentade variano da 54.0 a 56.4mm (min 52.0 max 60.0), mentre la media calcolata per l'intero campione è di 55.66mm (d.s. 1.98, n = 173). Nel corso del periodo di studio è possibile notare un certo incremento per entrambi i parametri nel periodo compreso tra le pentadi 50-
Le medie totali di queste due variabili presentano inoltre differenze significative con quelle calcolate nei mesi di ottobre-
I pesi medi calcolati per ciascuna pentade variano da 17.2 a 19.2g (min 14.9 max 24.8), mentre il peso medio totale calcolato per l'intero campione è di 18.64g (d.s. 1.70, n = 175), valore che non presenta differenza significativa con quello rilevato nei mesi di ottobre-
Balia nera Ficedula hypoleuca
Geonemia
Specie a corologia eurosibirica, la Balia nera presenta un'areale riproduttivo che va dall'Africa nord-

Fenologia della migrazione
Le catture sono state effettuate tra il 27 agosto ed il 3 ottobre 2004 (pentadi 48-
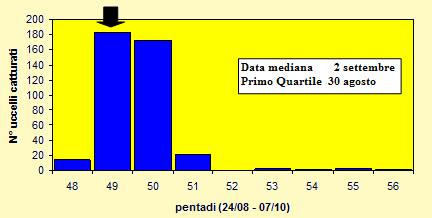
Figura 13. Andamento delle catture, nel periodo 27 agosto -
Come altri due migratori trans-
Analisi biometrica
In tabella 11 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
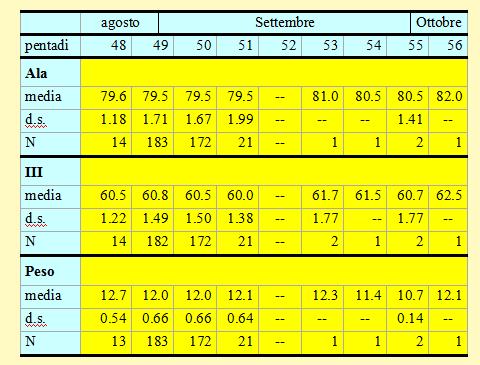
Tabella 11. Peso (in g) e misure biometriche (in mm) delle Balie nere catturate nell'Osservatorio ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 79.5 a 80.5mm (min 74.0 max 84.0), mentre la media calcolata per l'intero campione è di 79.52mm (d.s. 1.68, n = 395). Risulta evidente che nel periodo 27 agosto-
I pesi medi calcolati per ciascuna pentade variano da 10.7 a 12.7g (min 10.5 max 14.6), mentre il peso medio totale calcolato per l'intero campione è di 12.04g (d.s. 0.67, n = 394). Anche per il peso non si evidenzia nessuna marcata tendenza nel corso delle pentadi in cui è stato accertato il passaggio di un forte flusso migratorio; le differenze tra le medie non sono significative e, anche per i pochi soggetti inanellati nelle fasi finali della migrazione, non si riscontra nessun incremento delle condizioni fisiche in quanto il peso medio calcolato dall'8 settembre in poi risulta essere solo di 11.9g. Questi valori risultano decisamente inferiori rispetto alle medie riportate da Cramp e Perrins (1993) in autunno per la Francia meridionale (14.9g, d.s. 2.28, n = 129) e per la Spagna (adulti:14.5g, d.s. 1.92, n = 97; giovani: 13.6g, d.s. 1.65, n = 184) e sicuramente sono insufficienti per intraprendere voli su lunga distanza.
Analisi del grasso
I pesi medi corrispondenti a ciascuna classe di grasso sono riportati nella tabella 12. I valori rilevati nel corso di questa stagione confermano l'esistenza di una stretta correlazione tra il peso degli uccelli e la quantità di grasso accumulata stimata visualmente.
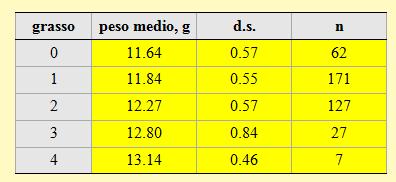
Tabella 12. Peso medio per ciascuna classe di grasso delle Balie nere catturate nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Se gli uccelli vengono classificati come magri (classi di grasso 0-
Come è stato accertato per il peso, anche per il grasso non si rileva nessun aumento significativo della frequenza di soggetti grassi nelle fasi terminali della migrazione.
Fringuello Fringilla coelebs
Geonemia
Specie a corologia olopaleartica, il Fringuello è comune ed ampiamente diffuso in tutta l'Europa fino alla Scandinavia con esclusione delle estreme latitudini settentrionali. Il comportamento migratorio presenta un'ampia variabilità di strategie con popolazioni parzialmente sedentarie ed altre totalmente migratrici; lo svernamento avviene prevalentemente all'interno dell'areale riproduttivo europeo, arrivando fino alle zone costiere dell'Africa settentrionale. In Lombardia è sedentario, migratore regolare e svernante, nidificante molto comune e diffuso in quasi tutti gli ambienti boschivi, distribuita in una fascia altimetrica che arriva fino a quasi 2000m.

Fenologia della migrazione
Le catture sono state effettuate tra il 28 agosto ed il 27 novembre 2004 (pentadi 48-
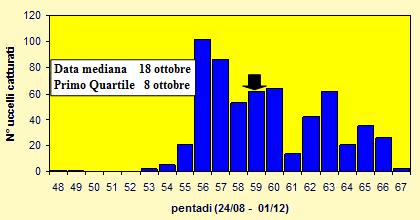
Figura 14. Andamento delle catture per pentade nel periodo 28 agosto-
Analisi biometrica
In tabella 13 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
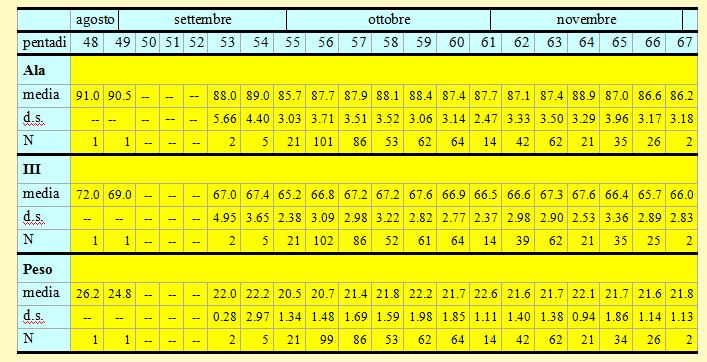
Tabella 13. Misure biometriche (in mm) e peso (in g) dei Fringuelli catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 85.7 a 89.0mm (min 79.0 max 95.5), mentre la media calcolata per l'intero campione è di 87.65mm (d.s. 3.45, n = 598). Le medie della III remigante primaria calcolate per ciascuna pentade variano da 65.2 a 67.6mm (min 60.0 max 74.0), mentre la media calcolata per l'intero campione è di 66.94mm (d.s. 2.98, n = 593). I pesi medi calcolati per ciascuna pentade variano da 20.5 a 22.6g (min 17.4 max 28.9), mentre il peso medio totale calcolato per l'intero campione è di 21.57g (d.s. 1.68, n = 595).
Le figure 15 e 16 mostrano le distribuzioni di frequenza, rispettivamente, delle misure della lunghezza dell'ala e del tarso.
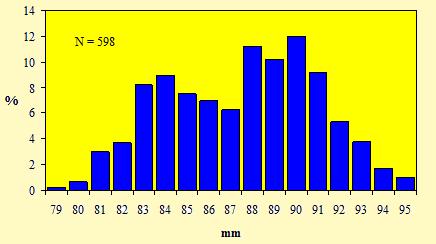
Figura 15. Distribuzione di frequenza delle misure della lunghezza dell'ala dei Fringuelli catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
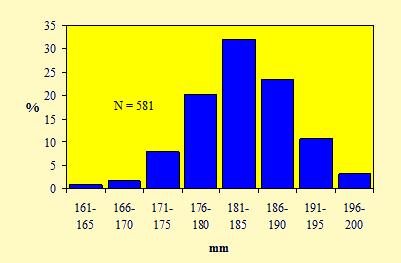
Figura 16. Distribuzione di frequenza delle misure della lunghezza del tarso dei Fringuelli catturati nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
L'andamento bimodale del grafico relativo alla lunghezza alare conferma l'esistenza di un dimorfismo dimensionale tra i sessi, con i maschi che risultano regolarmente più lunghi delle femmine. Diversamente, una chiara unimodalità emerge dalla figura 16, per cui la lunghezza del tarso non costituisce un carattere discriminante per determinare il sesso in questa specie. Inoltre questo andamento normale, cioè caratterizzato da una curva a campana con un unico picco, non permette di riconoscere nel campione, oltre che il sesso, anche l'eventuale presenza di individui appartenenti a due o più popolazioni distinte, originarie di regioni differenti.
Peppola Fringilla montifringilla
Geonemia
Specie a corologia eurosibirica, la Peppola è ampiamente diffusa dalla Scandinavia alla Siberia orientale; tutte le popolazioni sono migratrici e svernano quasi interamente al di fuori dei territori di riproduzione nell'Europa centrale e meridionale con notevoli fluttuazioni da un anno all'altro dovute soprattutto alla disponibilità di cibo. In Lombardia è migratrice regolare, svernante in numeri molto fluttuanti con occasionali segnalazioni nel settore alpino durante il periodo estivo.

Fenologia della migrazione
Le catture sono state effettuate tra il 5 ottobre ed il 26 novembre 2004 (pentadi 56-
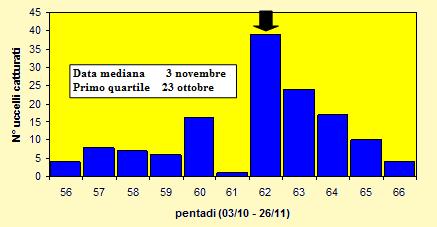
Figura 17. Andamento delle catture per pentade nel periodo 5 ottobre -
Analisi biometrica
In tabella 14 è riportata una sintesi delle variabili morfometriche e fisiologiche (lunghezza alare, III remigante primaria e peso) rilevate per ciascuna pentade.
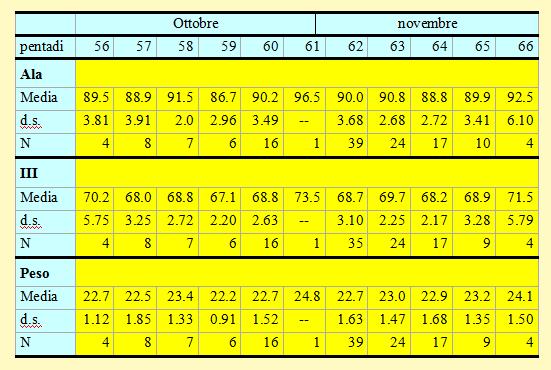
Tabella 14. Misure biometriche (in mm) e peso (in g) delle Peppole catturate nell'Osservatorio Ornitologico "Daniele Anesa", autunno 2004.
Le lunghezze alari medie calcolate per ciascuna pentade variano da 86.7 a 92.5mm (min 82.0 max 99.0), mentre la media calcolata per l'intero campione è di 89.96mm (d.s. 3.46, n = 136). Le medie della III remigante primaria calcolate per ciascuna pentade variano da 67.1 a 71.5mm (min 62.0 max 77.5), mentre la media calcolata per l'intero campione è di 68.90mm (d.s. 2.98, n = 131).
I pesi medi calcolati per ciascuna pentade variano da 22.2 a 24.1g (min 18.4 max 27.4), mentre il peso medio totale calcolato per l'intero campione è di 22.87g (d.s. 1.53, n = 135).
CONSIDERAZIONI CONCLUSIVE
Il motivo principale che ha spinto il G.I.L. ad organizzare l'Osservatorio Ornitologico "Daniele Anesa", oltre naturalmente allo studio della migrazione, è stato quello di mantenere vivo il Roccolo Ganda, un impianto datato intorno al 1750 e ritenuto "uno dei più belli della Lombardia", e la memoria del suo proprietario, l'Architetto Daniele Anesa che, con la sua passione e competenza era riuscito a creare una stazione ideale per lo studio delle migrazioni degli uccelli; si è inoltre voluto conservare la tradizione di tenerlo aperto a quanti desideravano visitarlo per apprezzarne in pieno tutto il suo fascino. Secondo quanto scrive il giornalista Ezio Pellegrini in un articolo pubblicato il 24 settembre sull'Eco di Bergamo questi obiettivi sono stati raggiunti.
Per quanto riguarda l'attività di cattura ed inanellamento, lo sforzo di campionamento è stato rilevante se si considera che il periodo stagionale ricoperto è stato molto esteso -
Occorre però rilevare che se si effettua un confronto con i risultati ottenuti negli anni precedenti ed in particolare con quelli a partire dal 1995, ne emerge un drastico calo del numero totale degli uccelli catturati. Sicuramente le sfavorevoli condizioni meteorologiche che si sono verificate dalla seconda metà di ottobre all'inizio di novembre, caratterizzate da un alternarsi continuo di perturbazioni, hanno avuto una pesante influenza in quanto la presenza di venti di forte intensità ha influito negativamente sia sul passaggio degli uccelli che sull'efficienza delle reti. Nel periodo succitato è stato infatti necessario chiudere più volte le reti per il vento e per la pioggia; se si considera che tale situazione è perdurata abbastanza a lungo, è probabile che movimenti migratori, per altro impossibili da monitorare, si siano comunque verificati.
Va comunque precisato che il numero delle catture è stato anche notevolmente condizionato dalla differente metodologia utilizzata (reti mist-
Per i Fringillidi in particolare si è verificata una marcata riduzione delle catture; l'esiguità dei campioni ottenuti potrebbe aver reso impreciso il monitoraggio dei flussi migratori che per qualche specie (Lucarino e Frosone) è stato addirittura impossibile. Se si considera che per Fringuello e Peppola è stata autorizzata la caccia in deroga consentendo il prelievo di piccole quantità, risulta evidente quanto sia invece importante ricavare dei trends attendibili sulle variazioni di densità delle popolazioni migranti, stabilire nel corso degli anni la percentuale di giovani presenti rispetto agli adulti e quindi conoscere l'eventuale impatto della caccia.
A questo proposito è importante ricordare che Schubert et al.
I dati raccolti nel corso del 2004 hanno consentito di determinare il rapporto giovani/adulti almeno per quelle specie di cui è stato possibile raccogliere un campione di una certa entità e potrebbe essere interessante il confronto con i risultati ottenuti da altre stazioni d'inanellamento lombarde. Molto importanti sono anche i dati relativi alla quantità di grasso presente nei migratori su lunga distanza; le determinazioni effettuate lasciano ipotizzare che la maggior parte degli individui preferisce adottare una strategia migratoria caratterizzata da soste intermedie, senza accumulare elevati depositi di grasso, prima di affrontare barriere particolarmente impegnative, quali il Mediterraneo ed il Sahara. Per i migratori su breve e media distanza è stato registrato che il periodo principale della migrazione avviene tra la fine di settembre e la fine di ottobre, i numeri degli individui osservati sono stati spesso anche molto consistenti ma la maggior parte degli uccelli tende a passare velocemente nell'area sorvolando l'impianto di cattura senza fermarsi.
In conclusione, si auspica che la Regione Lombardia e la Provincia di Bergamo, unitamente alla famiglia Anesa, confermino il loro interesse nel portare avanti questo progetto per ottenere un quadro completo dei movimenti migratori che avvengono attraverso le Prealpi bergamasche, per valutare nel corso degli anni la percentuale di giovani presenti rispetto agli adulti ed eventuali fluttuazioni delle popolazioni, per confrontare la qualità delle catture effettuate mediante reti mist-

Autunno al Roccolo Ganda
RINGRAZIAMENTI
L'eterogeneità di provenienza delle persone che hanno preso parte all'attività e le difficoltà di comunicazione e linguaggio sono state abilmente superate grazie alla collaborazione attiva di tutti, consentendo di creare un ottimo spirito di gruppo fondamentale per la buona riuscita della ricerca. Un doveroso ringraziamento va alla famiglia Anesa per aver messo a nostra disposizione il Roccolo Ganda e per averci consentito di proseguire il lavoro interrotto in quel triste febbraio 2003 ed in particolare a Silvia e Cristian per le ore trascorse insieme. Si ringrazia inoltre Vittorio Vigorita, U.O. Pianif. Faunistica e Venatoria Regione Lombardia per il suo impegno nel sostenere l'organizzazione dell'Osservatorio Ornitologico "Daniele Anesa". Ed infine l'ultimo pensiero non può che essere per Daniele, la cui assenza si è fatta sentire molto di più di quanto potessi immaginare sia sul piano pratico che su quello umano: è mancato il Direttore d'orchestra ma soprattutto è mancato l'Amico.
BIBLIOGRAFIA
Baker, K. (1993) Identification Guide to European Non-
Berthold, P. (1973) Proposals for the standardization of the presentation of annual events, especially of migrating data. Auspicium 5 (suppl.), 49-
Berthold, P. & Friedrich, W. (1979) Die Federlänge: Ein neues nützliches Flügelmass. Vogelwarte
Busse, P. (1976) The Spring Migration of Birds at the East Part of Polish Baltic Coast. Acta Zoologica Cracoviensia
Busse, P. & Maksalon, L. (1986) Biometrical variability of Song Thrushes migrating through Polish Baltic coast. Notatki Ornitologiczne
Cramp, S. (1988) The Birds of the Western Palearctic: Handbook of the Birds of Europe, the Middle East and the North Africa.
Cramp, S. & Perrins, C.M. (1993) The Birds of the Western Palearctic: Handbook of the Birds of Europe, the Middle East and the North Africa. Vol. VII. Oxford University Press.
Kaiser, A. (1993) A new multi-
Ioalé, P. & Benvenuti, S. (1982) Seasonal and diurnal variation of weight in four Passeriformes in autumn and winter. Avocetta
Lövei, G.L., Scebba, S., Minichiello, F. & Milone, M. (1984) Seasonal activity, wing shape, weights and fat reserve variation of Robins (Erithacus rubecula
Pennycuick, C.J. (1999) Measuring birds' wings for flight performance calculations (2nd Edition).
Scebba, S., Esposito, R. & Lövei, G.L. (1991) Distribuzione stagionale e variazioni del peso e del grasso nei migratori transahariani in un'isola del Mediterraneo. Atti VI Conv.ital. Ornit., Torino (1994): 449.
Scebba, S. (1993) Gli Uccelli della Campania. Ed. Esselibri, Napoli.
Scebba, S. e Moschetti, G. (2000) Biometria e rapporto giovani/adulti nel Tordo bottaccio Turdus philomelos: analisi preliminare della migrazione autunnale in Campania. Ucc. d'Italia 25: 18-
Schubert M., Fedrigo, A. & Massa, R. (1986) Timing and pattern of the post-
Svensson, L. (1992) Identification guide to European passerines.
SUMMARY
Roccolo Ganda (45°47'N 09°48'E), Aviatico, Bergamo (northern Italy) was founded about 1750 and its structure respects the traditional structure of the Roccoli in Bergamo's area. In particular its position is favourable to catch birds in migration. In fact the Roccolo Ganda is situated at the head of the Seriana Valley at the heigh of about 1100 metres along the route of birds migrating from the north-
A "Roccolo" is a traditional structure of the Bergamo area of Lombardy (North-
To capture birds in the Roccoli required skill, experience and a good knowledge of bird song and behaviour. These skills were traditionally passed from the one generation to the next and in Lombardy the skills used in each Roccolo were often slightly different from those used in the next. At the present time only a few Roccoli still operate and much of the traditional knowledge has been lost.
Roccolo Ganda began to work as a ringing station from October 1991 and a total of 97.371 birds of 87 species were ringed during 1991/2002. During the 2004 the research was supported by the Regione Lombardia and the Provincia of Bergamo and realized by the Gruppo Inanellamento Limicoli (G.I.L., Napoli). The main aims of project are:
-
-
-
-
-
The study has been based on continuous and standardized mist-
The research was supported in the fieldwork by the students of the University of Naples, volunteers from the Earthwatch Institute and Italian and foreign ornithologists.
Pubblicazione n° 58 del Gruppo Inanellamento Limicoli, Napoli