Sintesi resoconto 2014 - Gruppo Inanellamento Limicoli (GIL, Napoli)
Menu principale:
- Home Page
- L'Associazione
-
Ricerche in atto
- Migrazione autunnale Allodola
- Migrazione primaverile Tordo
- Ali Tordo bottaccio in Campania
- Stazione Monte Vaccaro
- Riserva Naturale Foce Volturno
- Inanellamento Medio Volturno
- Avifauna svernante
- Avifauna Foce Neto e Sila
-
Ricerche concluse
- Ali Turdidi e Beccacce ATC LT 2
- Stazione Malga Rosello
- Migrazione Caradriiformi
- Progetto Rondine
- Migrazione autunnale Tordo
- Ali Turdidi e Beccacce ATC CE
- Progetto Alauda 2000
- Osservatorio Roccolo Ganda
- Atlante Migrazioni Puglia
- Download
- News
Sintesi resoconto 2014
Studio sulla migrazione autunnale dell'Allodola in Campania: sintesi dei dati autunno 2014
Gruppo Inanellamento Limicoli (G.I.L., Napoli)
Federazione Italiana della Caccia -
Nell’autunno 2014, come negli anni precedenti, il monitoraggio della migrazione post-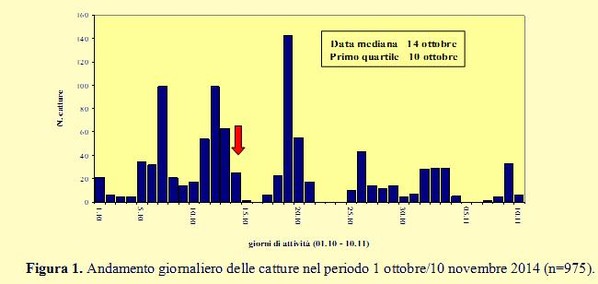
L’arrivo di una perturbazione caratterizzata da venti di forte intensità provenienti dai quadranti meridionali e da violenti temporali ha poi arrestato completamente la migrazione e non ha consentito di aprire l’impianto di reti il 5 ed il 6 novembre. Nei giorni successivi, con il migliorare delle condizioni meteo, sono stati osservati passaggi molto modesti ed anche il numero di individui inanellati è rimasto decisamente scarso fino alla fine del periodo di monitoraggio.
In base alla distribuzione temporale delle catture la data mediana (quando cioè è stato catturato il 50% di tutti gli uccelli), evidenziata nel grafico dalla freccia, è caduta il 14 ottobre, il 10 ottobre è terminato il primo quartile ed il 21 il terzo quartile: tra queste due date è risultato compreso il periodo principale della migrazione.
Anche quest’anno non vi sono state ricatture a distanza di uno o più giorni dall’inanellamento, per cui è possibile ritenere che la cattura non influenzi la strategia di migrazione e che gli uccelli non tendano a rimanere in prossimità dell’area di studio. Nella figura 2 la distribuzione delle catture è organizzata per decade. Come per lo scorso anno, la percentuale più alta è stata ottenuta nella seconda decade (48% con 468 inanellati) con un discreto valore anche nella prima (25,9% con 252 catture), mentre la terza decade ha fatto registrare la percentuale più bassa (12,3% con 120 inanellati).
Nella figura 2 la distribuzione delle catture è organizzata per decade. Come per lo scorso anno, la percentuale più alta è stata ottenuta nella seconda decade (48% con 468 inanellati) con un discreto valore anche nella prima (25,9% con 252 catture), mentre la terza decade ha fatto registrare la percentuale più bassa (12,3% con 120 inanellati).
Dalla tabella 1 si rileva infatti che i numeri delle catture ottenuti nelle quattro decadi di questa quinta stagione si discostano in modo evidente da quelli registrati negli anni precedenti con una riduzione particolarmente drastica nella terza decade di ottobre; ciò sta ad indicare che durante tutta la stagione il flusso migratorio è stato piuttosto modesto come conferma il totale di inanellati più basso tra quelli fin’ora ottenuti. Così nel 2014 in 38 giornate di attività si è raggiunta una quota media di 26 uccelli/giorno rispetto ad esempio al 2012 in cui 36 giornate hanno prodotto una quota di 58 uccelli ed al 2011 in cui in 37 giornate di attività si è avuta una quota di 39 catture.
OTT 1 |
OTT 2 |
OTT 3 |
NOV 1 |
TOT |
|
2006 |
421 |
566 |
494 |
81 |
1.562 |
2007 |
322 |
419 |
589 |
173 |
1.503 |
2010 |
34 |
201 |
1.142 |
193 |
1.570 |
2011 |
77 |
335 |
750 |
269 |
1.431 |
2012 |
399 |
1.169 |
300 |
231 |
2.099 |
2013 |
114 |
523 |
426 |
89 |
1.152 |
2014 |
252 |
468 |
120 |
135 |
975 |
1.619 |
3.681 |
3.821 |
1.171 |
10.292 |
|
Tabella 1. Numero di inanellati per decade (dati 2006 e 2007 da Scebba 2009). |
|||||

Anche se nei diversi anni è possibile osservare da una decade all’altra delle marcate fluttuazioni del numero di uccelli catturati, sul lungo periodo emerge che il maggior numero di inanellati è concentrato nella seconda e nella terza decade di ottobre, mentre nella prima decade di novembre le catture diminuiscono notevolmente.
Poiché negli anni 2006 e 2007 e 2010-
Anno |
n° catture |
mqrete x ore |
IC |
2006 |
1.562 |
73260 |
21,32 |
2007 |
1.503 |
72600 |
20,70 |
2010 |
1.570 |
73260 |
21,43 |
2011 |
1.454 |
68640 |
21,18 |
2012 |
2.099 |
70620 |
29,68 |
2013 |
1.152 |
65340 |
17,63 |
2014 |
975 |
69960 |
13,94 |
Tabella 2. Indice di Cattura (IC) per stagione |
|||
Uno degli obiettivi proposti dal “Management Plan for Skylark 2007–2009” della Commissione Europea (Petersen 2007) per una corretta gestione della specie è il monitoraggio della sex-
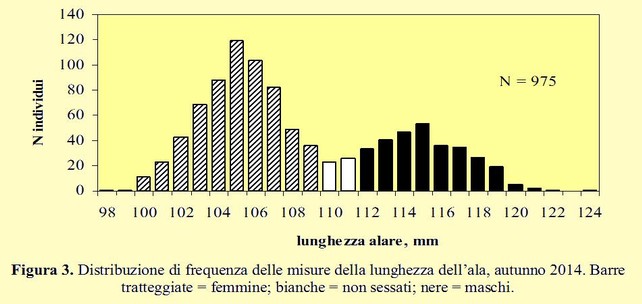
Come già osservato negli anni precedenti, la distribuzione dei dati relativi alla lunghezza dell’ala è molto ampia ed evidenzia una chiara bimodalità che va ascritta al dimorfismo sessuale; si può quindi ritenere che il campione rappresentato da individui di dimensioni più piccole sia costituito da femmine, mentre l’altro, che comprende soggetti di dimensioni maggiori, da maschi. Ciò risulta ancora più evidente se l’intero campione di dati raccolti dal 1998 al 2014 viene esaminato con il pacchetto statistico “mixtools” per R che permette di adattare una distribuzione bimodale ed ottenere la stima della media e della deviazione standard di ciascuna componente (Figura 4) (Scebba et al. 2015).
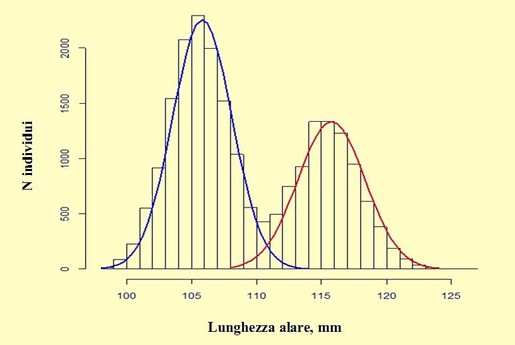
Figura 4. Distribuzione di frequenza delle misure della lunghezza dell’ala ottenuta con il pacchetto statistico “mixtools” per R, 1998-
Componente blu = femmine; componente rossa = maschi.
Dal grafico si rileva in modo evidente come l’andamento delle curve consenta di separare i due sessi in base alla lunghezza alare, mentre per il ristretto intervallo di sovrapposizione solo una modesta percentuale di individui rimane indeterminata ed un numero molto ridotto di uccelli risulta non correttamente sessato. Dall’analisi statistica è emerso che la lunghezza alare media è di 105,83mm (d.s. 2,285) per le femmine e di 115,76mm (d.s. 2,583) per i maschi, valori simili a quelli ottenuti con il metodo di Harding-
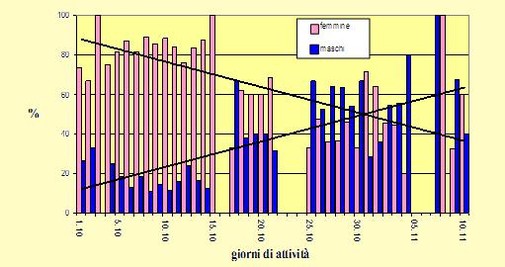
Figura 5. Fenologia della migrazione autunnale basata sulla percentuale di maschi e femmine nel 2014 con linee di tendenza.
Questa progressiva diminuzione delle femmine nel corso della migrazione, regolarmente evidenziata in tutti gli anni in cui è stato condotto il monitoraggio (1998-
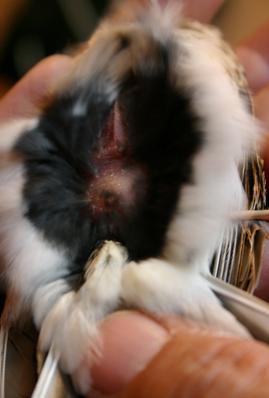
Gli aspetti energetici legati alla migrazione dell’Allodola non sono mai stati studiati in dettaglio, nonostante l’entità dell’accumulo di grasso sottocutaneo rappresenti un parametro importante per valutare lo stato dell’animale in quanto il grasso fornisce l’energia necessaria per sostenere la migrazione.
Nel corso di questi anni sono stati rilevati peso corporeo e grasso su più di 20.000 soggetti. Questi dati possono essere usati per studiare i diversi aspetti della migrazione, in particolare per determinare le strategie adottate in termini di rotta e di fenologia e per valutare la variazione inter-
Autunno 2010 |
Autunno 2011 |
Autunno 2012 |
Autunno 2013 |
Autunno 2014 |
||||||
Classi di |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
0 |
2 |
5 |
10 |
14 |
2 |
9 |
11 |
34 |
2 |
10 |
1 - |
162 |
297 |
86 |
203 |
96 |
370 |
145 |
391 |
49 |
250 |
3 - |
388 |
524 |
331 |
445 |
232 |
602 |
191 |
276 |
205 |
332 |
5 - |
61 |
55 |
169 |
126 |
36 |
65 |
28 |
27 |
40 |
31 |
Totale |
613 |
881 |
596 |
788 |
366 |
1.046 |
375 |
728 |
296 |
623 |
Tabella 3. Distribuzione (N e %) delle allodole nelle diverse classi di grasso, separate per sesso. |
||||||||||
Nel 2014, come in quasi tutti gli altri anni, la percentuale di allodole che non hanno presentato depositi di grasso visibili (classe 0) è risultata molto bassa, come anche le percentuali di soggetti inclusi nelle classi 1 e 2, cioè con depositi di grasso molto ridotti. Risulta invece elevato il numero di uccelli inclusi nelle classi di grasso 3-
Nel corso dell’autunno del 2014 è stata riscontrata una marcata diminuzione del flusso migratorio che ha portato ad una notevole riduzione del numero delle catture. Tra il 1 ottobre ed il 10 novembre con 38 giornate effettive di attività sono stati inanellati soltanto 975 uccelli (tabella 1), uno dei risultati più bassi fin’ora ottenuti, in quanto solo il 2003 è stato peggiore con 962 inanellati. Questa flessione, non influenzata dalle condizioni meteorologiche che si sono mantenute abbastanza favorevoli per gran parte del periodo, può rientrare nel normale andamento della fenologia della migrazione in quanto, da un anno all'altro, spesso si verificano delle significative fluttuazioni nei passaggi dei migratori, come descritto per questa specie già da Duse e Toschi (1930).
Bibliografia
Duse A. & Toschi A. (1930) Contributo allo studio delle migrazioni dell’Allodola, del Tordo e del Fringuello. Ricerche Zoologia applicata alla Caccia 1, pp. 23.
Harding J.P. (1949) The use of probability paper for the graphical analysis of polymodal frequency distributions. Journal of the Marine Biological Association of the United Kingdom 28: 141-
Kaiser A. (1993) A new multi-
Petersen B.S. (2007) Management Plan for Skylark (Alauda arvensis) 2007–2009.
Scebba S. (2009) Rapporto “Alauda 2000”. Indagine sulla migrazione autunnale nella Piana del Volturno (Caserta) in base all’attività di inanellamento: autunno 1998 – 2007. Studio Lito Clap, Bergamo. 125 pp.
Scebba S., Soprano M. & Sorrenti M. (2015) Sex-