Sintesi resoconto 2012 - Gruppo Inanellamento Limicoli (GIL, Napoli)
Menu principale:
- Home Page
- L'Associazione
-
Ricerche in atto
- Migrazione autunnale Allodola
- Migrazione primaverile Tordo
- Ali Tordo bottaccio in Campania
- Stazione Monte Vaccaro
- Riserva Naturale Foce Volturno
- Inanellamento Medio Volturno
- Avifauna svernante
- Avifauna Foce Neto e Sila
-
Ricerche concluse
- Ali Turdidi e Beccacce ATC LT 2
- Stazione Malga Rosello
- Migrazione Caradriiformi
- Progetto Rondine
- Migrazione autunnale Tordo
- Ali Turdidi e Beccacce ATC CE
- Progetto Alauda 2000
- Osservatorio Roccolo Ganda
- Atlante Migrazioni Puglia
- Download
- News
Sintesi resoconto 2012
Studio sulla migrazione autunnale dell'Allodola in Campania: sintesi dei dati autunno 2012
Gruppo Inanellamento Limicoli (G.I.L., Napoli)
Federazione Italiana della Caccia -
Anche per l'autunno 2012 è stato effettuato il monitoraggio della migrazione post-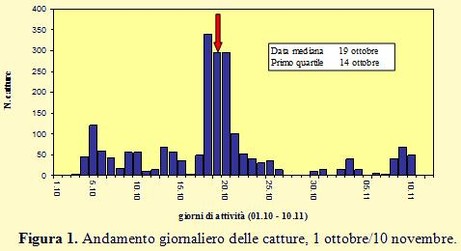 Nei giorni successivi le catture si sono poi ridotte rapidamente, in particolare per l’arrivo di una serie di perturbazioni caratterizzate da intense precipitazioni e fortissimi venti dai quadranti meridionali. Successivamente, quando le condizioni meteorologiche sono migliorate e, soprattutto, è scemata l’intensità del vento, è stata ancora registrata una modesta ondata di migratori, così come già era stato osservato in passato, che ha consentito di inanellare 156 uccelli negli ultimi tre giorni della prima decade di novembre e che hanno rappresentato la coda della migrazione autunnale. La data mediana è caduta il 19 ottobre, quattro giorni prima rispetto al 2010 e ben nove rispetto al 2011, il primo quartile è terminato il 14 ottobre ed il terzo quartile il 21. Tra queste due date risulta compreso il periodo principale della migrazione.
Nei giorni successivi le catture si sono poi ridotte rapidamente, in particolare per l’arrivo di una serie di perturbazioni caratterizzate da intense precipitazioni e fortissimi venti dai quadranti meridionali. Successivamente, quando le condizioni meteorologiche sono migliorate e, soprattutto, è scemata l’intensità del vento, è stata ancora registrata una modesta ondata di migratori, così come già era stato osservato in passato, che ha consentito di inanellare 156 uccelli negli ultimi tre giorni della prima decade di novembre e che hanno rappresentato la coda della migrazione autunnale. La data mediana è caduta il 19 ottobre, quattro giorni prima rispetto al 2010 e ben nove rispetto al 2011, il primo quartile è terminato il 14 ottobre ed il terzo quartile il 21. Tra queste due date risulta compreso il periodo principale della migrazione.
Nella figura 2 le catture sono organizzate per decadi; l’andamento si discosta in modo evidente da quelli registrati negli anni 2010 e 2011 che, invece, sono risultati abbastanza simili, anche se con percentuali differenti.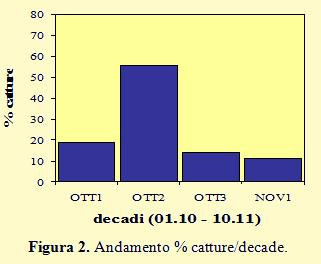
Nel 2010 e nel 2011 i valori più elevati sono stati infatti riscontrati sempre nella terza decade di ottobre, rispettivamente con il 73% (1.142 inanellati) e con il 51,7% (750 inanellati), mentre nel 2012 la percentuale più alta di catture si è avuta nella seconda decade (55,7% con 1.169 inanellati) ma intensi movimenti migratori sono avvenuti anche nella prima (19%). Se si confrontano questi dati con quelli riportati nel Progetto “Alauda 2000” per gli anni 2006 e 2007, in cui il monitoraggio della migrazione è stato condotto nella stessa area di cattura e con un transetto di reti verticali di tipo mist-
OTT 1 |
OTT 2 |
OTT 3 |
NOV 1 |
TOT |
|
2006 |
421 |
566 |
494 |
81 |
1.562 |
2007 |
322 |
419 |
589 |
173 |
1.503 |
2010 |
34 |
201 |
1.142 |
193 |
1.570 |
2011 |
77 |
335 |
750 |
269 |
1.431 |
2012 |
399 |
1.169 |
300 |
231 |
2.099 |
Tabella 1. Numero di inanellati per decade (dati 2006 e 2007 da "Alauda 2000"). |
|||||
Poiché il monitoraggio della migrazione è stato condotto nell’autunno del 2006 e del 2007 e quindi negli anni 2010, 2011 e 2012 con una metodologia di cattura completamente standardizzata, è possibile valutare con maggior precisione l’andamento dei passaggi avvenuti nelle cinque stagioni mediante un indice di cattura (IC). È stata quindi considerata tutta l’attività espletata, dalla prima sessione di cattura effettuata il 1 ottobre fino all’ultima avvenuta il 10 novembre, in modo da coprire l’intero periodo della migrazione (tabella 2). L’indice di cattura è stato ricavato dividendo il numero degli uccelli catturati in una determinata stagione per il prodotto del numero di metri quadrati di rete utilizzati moltiplicato il numero di ore di esposizione delle stesse e moltiplicando poi il risultato finale per 1000.
Anno |
n° catture |
mq x ore |
IC |
Dall’analisi dei dati riportati in tabella si rileva un indice di cattura che può essere considerato costante nei primi quattro anni mentre nel 2012 vi è un forte incremento dovuto agli intensi passaggi osservati tra il 17 ed il 21 ottobre, probabilmente favoriti da venti provenienti dai quadranti settntrionali. Va infatti osservato che condizioni climatiche favorevoli o, al contrario, la presenza di venti molto forti anche in una sola giornata di intensi passaggi, possono decisamente influenzare il numero delle catture (soprattutto per le condizioni delle reti) e questo spiegherebbe anche le modeste differenze che si riscontrano da un anno all’altro. |
||||||
2006 |
1.562 |
73260 |
21,32 |
|||||||
2007 |
1.503 |
72600 |
20,70 |
|||||||
2010 |
1.570 |
73260 |
21,43 |
|||||||
2011 |
1.454 |
68640 |
21,18 |
|||||||
2012 |
2.099 |
70620 |
29,68 |
|||||||
Tabella 2. Attività di cattura durante la migrazione post- |
||||||||||
Come richiesto dal "Management Plan for Skylark" della Commissione Europea (Petersen 2007) è anche proseguito l'analisi della sex-
Va sottolineato che per la determinazione del sesso sono stati utilizzati i parametri riportati nel Progetto “Alauda 2000” che coincidono con quelli usati in Francia per distinguere i due sessi (Demongin 2006) ed in particolare dal programma di ricerca “Alouette” condotto dal “Centre de Recherches sur la Biologie des Populations d’Oiseaux” (Centro d’inanellamento francese) che coinvolge una trentina di inanellatori i quali tra il 1989 ed il 2000 hanno inanellato più di 50.000 uccelli (Teulière 2003). Nella figura 3 viene riportata la fenologia della migrazione basata sulle percentuali di maschi e femmine catturati, inanellati e sessati nel 2012 lungo la costa sud-
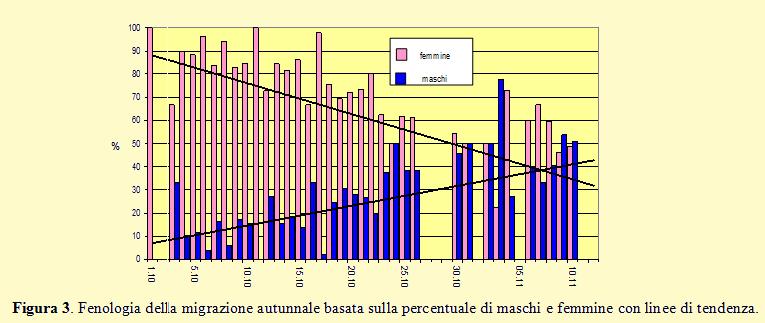
Dal grafico si osserva una maggiore presenza di femmine dalle prime fasi della migrazione fino alla fine della seconda decade di ottobre, quando poi inizia un forte passaggio caratterizzato da un elevato numero di maschi che rimangono numerosi per tutto il successivo periodo. Se la sex-
Questa progressiva diminuzione delle femmine nel corso della migrazione è determinata dalla tendenza dei maschi a migrare più tardi e ad allontanarsi di meno dalla aree di riproduzione; è poi probabile che le variazioni nel numero di femmine presenti nel flusso migratorio autunnale che si verificano da un anno all’altro possano essere dovute alle condizioni meteorologiche e/o alle disponibilità alimentari.
Nell'ambito del progetto di ricerca l’esame delle condizioni fisiche degli uccelli in migrazione ha un'importanza rilevante ed è effettuato valutando l’entità dei depositi di grasso sottocutaneo utilizzati come “carburante” durante il viaggio. Ciò può consentire di determinare l’autonomia potenziale dei migratori e può fornire informazioni utili per determinare le strategie di migrazione adottate. Nella tabella 3 vengono riportate le percentuali ed il numero di individui, separati per sesso, inclusi nelle diverse classi di grasso, calcolate nei tre anni di studio. Le percentuali di allodole che non presentano depositi di grasso visibili (classe 0) o, comunque, molto ridotti (classi 1 e 2) risultano sempre molto basse in quanto va ricordato che il monitoraggio viene effettuato su uccelli in piena attività migratoria. Va poi osservato che per entrambi i sessi si rileva un elevato numero di uccelli con abbondanti depositi adiposi dovuto probabilmente al passaggio di una nutrita componente di soggetti in migrazione verso aree di svernamento più meridionali. È interessante osservare che le femmine presentano sempre depositi adiposi più ridotti rispetto a quelli dei maschi mostrando percentuali più elevate nelle classi di grasso 0-
Tabella 3. Distribuzione delle allodole nelle classi di grasso, separate per sesso. |
||||||
Autunno 2010 |
Autunno 2011 |
Autunno 2012 |
||||
Classi di |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
♂♂ |
♀♀ |
0 |
2 |
5 |
10 |
14 |
2 |
9 |
1 - |
162 |
297 |
86 |
203 |
96 |
370 |
3 - |
388 |
524 |
331 |
445 |
232 |
602 |
5 - |
61 |
55 |
169 |
126 |
36 |
65 |
Totale |
613 |
881 |
596 |
788 |
366 |
1.046 |
Se si considera l’intero campione ottenuto per anno si rileva che le percentuali di uccelli con scarse riserve di grasso (classi 0, 1 e 2) non mostrano variazioni importanti (30,9% nel 2010, 22,7% nel 2011 e 34,1% nel 2012); inoltre quelli che mostrano un punteggio di grasso uguale o superiore a 3 posseggono energie sufficienti per proseguire la migrazione senza effettuare soste e sono in grado di raggiungere zone di svernamento poste più a sud dell’area di studio.
E' evidente che i dati raccolti nell'autunno 2012 costituiscono un'importante fonte di informazioni ed arricchiscono la banca-
BIBLIOGRAFIA
Demongin, L. (2006) Guide d’identification en main des oiseaux d’Europe. Identification, biométrie, mue, sexe, âge. Provisional version, unpublished, 164 pp.
Petersen, B.S. (2007) Management Plan for Skylark (Alauda arvensis) 2007–2009. Office for Official Publications of the European Communities, Luxembourg.
Scebba, S. (2009) Rapporto “Alauda 2000”. Indagine sulla migrazione autunnale nella Piana del Volturno (Caserta) in base all’attività di inanellamento: autunno 1998 – 2007. Studio Lito Clap, Bergamo.
Schekkerman, H. (1999) Sex bias and seasonal patterns in tape-
Teulière, J.M. (2003) Baguage de l'Alouette des champs (Alauda arvensis
Vignes, J.C. (2001) Observations sur les migrations et la biométrie de l’Alouette des Champs (Alauda arvensis